
In the mid-nineteenth century, the concept of muscle behaving like a stretched spring was developed. This elastic model of contraction predicted that the energy available to perform work was established at the start of a contraction. Despite several studies showing evidence inconsistent with the elastic model, it persisted into the twentieth century. In 1923, W. O. Fenn published a paper in which he presented evidence that appeared to clearly refute the elastic model. Fenn showed that when a muscle performs work it produces more heat than when contracting isometrically. He proposed that energy for performing work was only made available in a muscle as and when that work was performed. However, his ideas were not adopted and it was only after 15 years of technical developments that in 1938 A. V. Hill performed experiments that conclusively disproved the elastic model and supported Fenn’s conclusions. Hill showed that the rate of heat production increased as a muscle made the transition from isometric to working contraction. Understanding the basis of the phenomenon observed by Fenn and Hill required another 40 years in which the processes that generate force and work in muscle and the associated scheme of biochemical reactions were established. Demonstration of the biochemical equivalent of Hill’s observations—changes in rate of ATP splitting when performing work—in 1999 was possible through further technical advances. The concept that the energy, from ATP splitting, required to perform work is dynamically modulated in accord with the loads a muscle encounters when contracting is key to understanding muscle energetics.
Avoid common mistakes on your manuscript.
The key concept in muscle energetics that is examined in this article is that muscle energy expenditure is determined by the mechanical loading a muscle encounters as it contracts. From the mid-nineteenth century until the 1930s, a contrasting idea held sway. Then it was thought that muscle energy expenditure was determined simply by the length of a muscle at the start of a contraction and that the mechanical behaviour of active muscle, and its associated energetics, could be understood by considering the muscle to act like a stretched spring. This was termed the elastic theory of contraction. Within that period, in 1923, a 30-year old American, Wallace O. Fenn, on a travelling fellowship to the UK wrote a paper in which he presented evidence that vigorously challenged the elastic theory. Fenn’s work might have been expected to overturn the elastic theories but that was not the case. Instead, the elastic theory continued to be used and elaborated. In this article, Fenn’s work is described in the context of the foundational work of European scientists in the nineteenth century and the prolonged pursuit of conclusive evidence about the validity or otherwise of the elastic theory by A. V. Hill in the twentieth century.
Central to this field of research was the use of measurements of the heat produced by muscles when they contract to examine fundamental aspects of muscle contraction. The application of physics to the study of biology in the nineteenth century, in particular knowledge of the behaviour of elastic materials and developing ideas about conservation of energy, provided both the impetus to explore muscle heat production in relation to activity and the development of a framework within which muscle heat output could be understood. The ultimate rejection of those ideas opened the field for a new framework to encompass the production of heat by active muscle and the processes underlying contraction.
There are recent comprehensive reviews covering biochemical aspects of energetics and the relationship between muscle heat production and biochemical changes (Barclay 2015a; Barclay and Loiselle 2021; Kushmerick 1983; Woledge et al. 1985) and on muscle biochemistry during exercise (e.g. Brooks 2012). It is not intended that this review overlap any more than necessary with such work. Instead, the current work will focus on the role that measurements of muscle heat output have played in the evolution of one of the key concepts in muscle energetics. Heat measurements are complicated and many of the technical minutiae that are often prominent in papers in the area are of somewhat esoteric interest. Consequently, it is not intended to focus on those aspects but rather to attempt to give a broad view of how, in this particular case, advances in physiological knowledge were slowed by the time taken to both overcome embedded ideas and to make necessary technical advances.
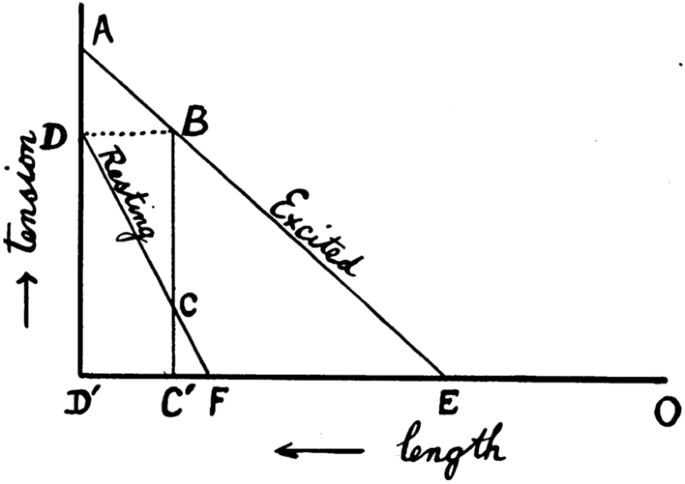
The modern approach to the study of muscle physiology arose in Europe in the 1830s and 1840s (for reviews of work in this and earlier eras, see Hill 1959, 1965b; Needham 1971). An important step was Theodor Scwhann’s 1835 demonstration that an isolated frog muscle produced its greatest force upon stimulation when close to its in vivo length and that less force was produced when the muscle was held at shorter lengths. That observation prompted Johannes Muller (Berlin, Germany) to propose that muscle be considered as an elastic body, with its force output reflecting its extension (or length). Eduard Weber (Liepzig, Germany) extended that idea by stating that the elastic properties of muscle altered upon stimulation so that the stimulated muscle behaved as a “new elastic body”, that is, with different elastic properties to an unstimulated muscle. Muller and Weber compared a stimulated muscle to a stretched spring and as such the muscle contained elastic potential energy (Fig. 1). If the muscle lifted a load, and in so doing shortened, then some of the elastic energy would be converted into mechanical work. Any elastic energy not converted into work would be dissipated as heat when the muscle force declined during relaxation. In this model, the only determinant of the amount of energy liberated as heat and work was the length of the muscle at the time it was stimulated. This simple phenomenological model of muscle mechanics, and variations of it, was to persist for almost 100 years.
The idea that the elastic energy would appear as either work or heat arises from the idea of conservation of energy which was a subject of much interest in European science at the time. Hermann Helmholtz (Berlin, Germany) had been seeking examples supporting the principle of energy conservation and one of these related to muscle. Increases in muscle temperature during activity had been recorded in humans and animals (Becquerel and Breschet 1835) and (Liebig 1842) proposed that the heat produced by muscle arose from chemical changes occurring within the muscle. Helmholtz recognized that the warming of muscles could occur by the transfer of heat to them from circulating blood. To address that possibility, he measured the contraction-induced change in temperature of a frog muscle preparation isolated from its blood supply and stimulated via its nerve (Helmholtz 1848). Helmholtz measured the temperature change using a thermopile and a galvanometer to measure the current flowing through the thermopile. The thermopile consisted of three pairs of thermocouples in series, with one of each pair (the “active” thermocouple) embedded in the stimulated muscle and the other (the “reference” thermocouple) embedded in the unstimulated contralateral muscle. This system could easily resolve the differences in temperature between the two muscles (up to 0.2 ºC) that occurred in the muscle in response to tetanic stimulation. Helmholtz’s method of using a thermopile to measure changes in muscle temperature, and thus to potentially calculate the amount of heat produced by the muscle, paved the way for subsequent workers to address the validity of the elastic model.
Within the framework of the elastic theory, it was thought that as force developed in an isometric contraction, heat was produced from the, presumably chemical, processes that accounted for the development of the elastic potential energy (Hill 1913a; Hill and Hartree 1920). If the muscle performed no work, as in an isometric contraction, then when force declined during relaxation an amount of heat equivalent to the elastic potential energy would be produced and the total heat output would be the sum of the heat from process creating the elastic energy and the heat from the dissipation of the elastic energy. If the muscle performed work during the contraction, some of the potential energy would be converted to work, and less heat would arise from the decay of elastic potential energy. Therefore, if the elastic theory was correct, less heat would be produced in a working contraction than in an isometric contraction but the total energy output (i.e. heat + work) would be the same. The predictions of the elastic model in term of heat output and total energy output are summarised in Table 1.
Hill thought that the heat measurements made by Heidenhain and Fick were insufficiently reliable to provide evidence strong enough to reject the elastic theory. Hill carried out several studies that included examination of aspects of the elastic theory (Evans and Hill 1914; Hill 1913a; Hill and Hartree 1920). From his evidence, Hill agreed with Blix that isometric contractions produced more heat than those with shortening. However, these experiments were unconvincing, often involving manipulation of multiple, most likely interacting, variables and comparisons which were unlikely to distinguish between elastic or alternative mechanisms. In retrospect, Hill stated that a major impediment in these studies was that ideas of what was expected were unclear (Hill 1965b). In a positive step, Hill had settled on using the frog sartorius muscle because it had straight fibres and was thin. The straight fibres provide accurate measurements of muscle work output and the thinness provided rapid thermal equilibrium with the thermopile and reduced limitations to diffusive O2 supply throughout the muscle cross section (Hill 1928).
In 1922 and 1923, Wallace O. Fenn, an American research fellow working in A. V. Hill’s laboratory in the UK, carried out an extensive series of experiments to test the idea that energy output as work and heat depended only on initial muscle length. Hill suggested these experiments to Fenn and provided the equipment (Fenn 1923). The Introduction to Fenn’s (1923) paper is evidence that Hill and Fenn had clarified what would be expected if the elastic theory was correct and Fenn (1923) summarised the view of adherents of the elastic theory thus,
The amount of elastic potential energy which could be recovered as work … had no relation to the total energy liberated.
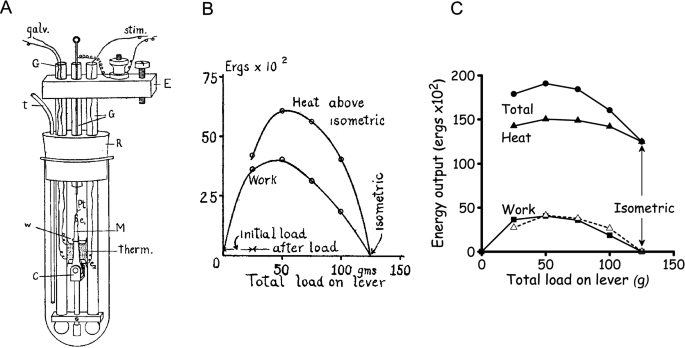
Fenn’s experiments were designed to determine the relationship, if one existed, between the total energy liberated, as work and heat, and the amount of work performed. This clarity of purpose was a major advance of Fenn’s work over the earlier studies. Furthermore, his experiments were more systematic than much of the earlier work. Fenn used Hill’s latest muscle heat measuring system (Hill and Hartree 1920) (Fig. 3A). The heat recordings were still quite slow, compared to the time course of muscle force production. For example, with frog muscle at close to 0 ºC, twitch contractions, which were commonly used at that time, were complete in ~ 0.5 s. In response to a heating pulse of similar duration, the galvanometer output took ~ 5 s to reach a peak. So, again, the records gave no information about the time course of heat production; instead, only the cumulative heat produced up to a time some seconds after a contraction was measured (Hill 1965b). Consequently, the differences in heat production between isometric and working contractions had to be determined by subtraction of two large and similar quantities. Many years later, it was also discovered that, despite Hill and Fenn’s best efforts, there was a substantial heat calibration error; the calculated heat values were ~ 1.5-times too large (Hill and Woledge 1962) but this does not affect the conclusions.
Fenn employed both a traditional lever system to perform afterloaded isotonic contractions and also an “inertia lever”, based on a device used by Fick (Hill 1920). With an isotonic lever, the muscle raised the load when its force generating capacity exceeded the load and then it lowered the load during relaxation. When the load was lowered an extra amount of heat, equivalent in magnitude to the work done, was produced. The total heat produced was then the sum of the heat generated by the muscle and the thermal equivalent of the work done. Hill, however, was not convinced that all the heat produced from lowering the load could be measured. The inertia lever allowed the muscle to relax against a very low load so that the heat output was measured directly rather than by subtraction from the total energy output as with the isotonic lever. Fenn’s textual clarity (see next paragraph) did not always extend to the tabulated data: confusingly, in his papers Fenn variously labels as “heat” that measured using the inertia lever and also that from afterloaded contractions which included the heat produced when the load was lowered (thus really total energy output, heat + work), making some of the tabulated data difficult to understand (Hill 1965b).
Fenn published his results in two papers (Fenn 1923; 1924), with the most important results in the first of these. The papers are notable not only for their scientific contribution but also for the clarity with which they are written, with clear expression of ideas underlying the experiments and lucid conclusions. It was also notable that Fenn used graphs to illustrate his results at a time when this was not common (tabulated data were more common). Fenn consistently observed that the total energy output was not independent of load; rather, more energy was liberated from muscles when they performed work than in an isometric contraction (Figs. 3B and C). Additionally, the extra energy was not just the work performed, additional heat was also produced. He found similar results for twitches and short tetani and using afterloaded isotonic contractions and the inertia lever. Fenn (1923) gave a concise conclusion.
Whenever a muscle shortens upon stimulation and does work in lifting a weight, an extra amount of energy is mobilised which does not appear in an isometric contraction. Hence less energy is liberated in an isometric contraction than in any contraction in which the muscle is allowed to shorten.
A striking aspect of the study from a modern perspective is that there was no form of statistical analysis available; the design of the experiments certainly lends them to multivariate analysis. In the second paper Fenn addressed the implications of his results for ideas about how muscle works, explicitly rejecting the elastic theory. Fenn stated that although a stimulated muscle might produce considerable tension, it may still possess little elastic energy, and thus the energy required to shorten and perform work had to be liberated as the shortening proceeds (Fenn 1924). Fenn gave full acknowledgement to the nineteenth century European researchers, particularly Fick, who had come to the same conclusion, albeit with less convincing experimental evidence.
In 1923, Hill was awarded the Nobel Prize for the previous year and in his Nobel speech, given shortly before publication of Fenn’s 1923 paper, he briefly described Fenn’s results. He concluded,
We are studying here the curious power which a muscle possesses of adapting its liberation of energy to the work it has to do. The subject is new and results are only now beginning to appear, but it is certain that we are dealing with a very fundamental property of the muscular machine.
Despite those sentiments, Hill and those working in his laboratory not only continued to use the elastic model as a framework for interpreting experiments but further developed it, incorporating a viscous element (Hill 1922). The new idea was that the solution within muscle cells had to be forced through some sort of structural network when the muscle shortened and that this provided a resistance to shortening, giving rise to a system with mechanical behaviour analogous to that of a damped spring. Overcoming the viscous resistance required some of the elastic potential energy which was converted into heat rather than work. The viscous-elastic model could account quantitatively for a range of phenomena (e.g. effects of shortening on force and work output and why measured efficiency was lower than that predicted from the supposed elastic potential energy) in both human (Hill 1922) and isolated frog muscles (Gasser and Hill 1924). Levin and Wyman (1927) further elaborated the model to account for their observations on “tension-length curves” (which would now be described as “work loops”) to consist of both a damped elasticity (i.e. the component affected by muscle viscosity) in series with an undamped elasticity. This model could account for the curved work-shortening velocity curves they measured with muscles from many different animals. These curves were actually the same as force–velocity curves and were the first of these to be measured.
Experiments designed to repeat Fenn’s work were also carried out. Jeffries Wyman (1926), a doctoral student working in Hill’s lab and using a new inertia lever design (Fig. 4A), found heat production by tortoise muscle (used because its slow contraction and heat production kinetics might compensate for the slow response of the heat recording apparatus) was greater in a contraction with shortening than in an isometric contraction (Fig. 4B) but less in a contraction with stretching than one with shortening. These results were similar to those of Fenn (1924) and Fick (1881). Whereas those authors had concluded that the results indicated that physiological processes accounted for the heat and work output, Wyman produced what now appear rather contrived arguments—a variable fraction of potential elastic energy not used as work was reabsorbed during relaxation rather than appearing as heat—to support the idea that all the thermal changes could be explained by processes that were physical (i.e. dissipation of stored potential energy and effects of viscosity) rather than physiological. In 1928, Hill and Hartree repeated Fenn’s experiments on frog muscle and surprisingly found that whereas they could replicate Fenn’s results for tetanic contractions they could not replicate his observation of extra heat in twitches (Hartree and Hill 1928a).
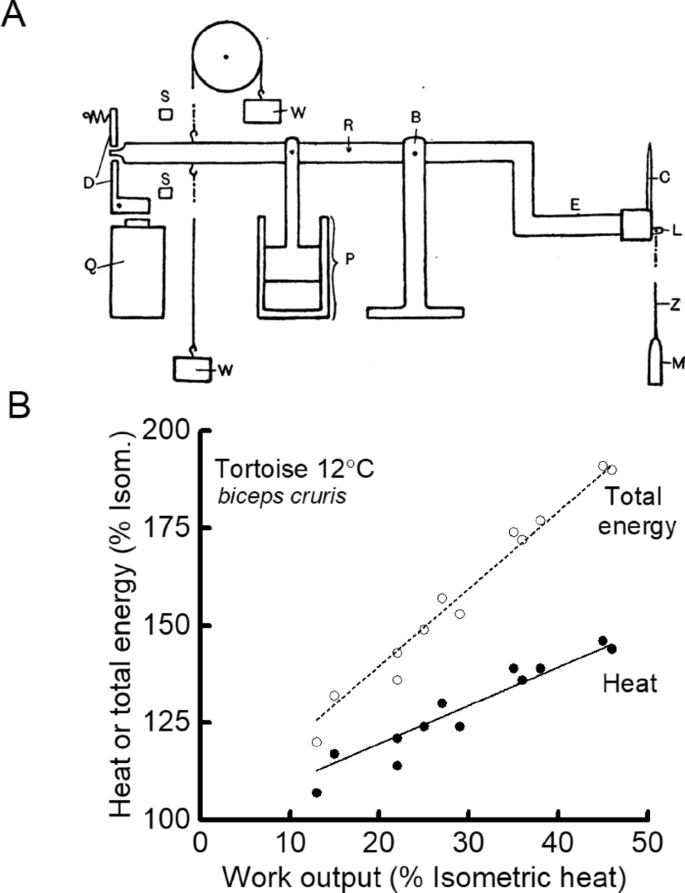
In his 1965 book, Hill lists 10 papers published between 1925 and 1932 that had attempted to repeat or expand upon Fenn’s results but with little success. Hill summarised the post-Fenn attempts to confirm the 1923 results.
In Hill (1925) “a possible mechanism of the Fenn effect” was discussed, but this lead nowhere. In 1925 Hartree attempted to analyse the course of the heat production during a contraction with shortening and work; but the results are doubtful because of the absence of a protecting region in the thermopile, and he did not realise how much work was done in an “isometric” contraction. In Hartree and Hill (1928b) the “Fenn effect” was clearly confirmed in tetanic contractions but not in twitches; the failure to do so was probably due to an error in the heat calibration. And so on.
Despite Hill’s rather pessimistic view (in retrospect, he felt that a great amount of work had been wasted due to his own inability to discard the elastic model), there was now a considerable body of information indicating that, even accepting the technical limitations, it was unlikely that the predictions of the elastic or viscous elastic model were correct. Furthermore, in successive heat studies from 1925 to 1928, Hill and his colleagues had made important developments of the heat measuring equipment and methods of analysis of heat records and these would eventually contribute to the system that was to settle the debate as to whether the performance of work was powered by energy from previously stored elastic energy or by energy from chemical reactions linked to the action of doing work.
In the decade after the Hartree and Hill (1928a) paper, Hill’s attention moved away from what he had termed “the Fenn effect” (Hill 1925). In 1935, Fenn returned to the discussion, publishing a provocative paper in which he produced an independent line of evidence to further support his contention that the elastic theories were untenable. Fenn and Marsh (1935) proposed an experimental test based on a viscous elastic model of muscle mechanical behaviour suggested by Levin and Wyman (1927). That model consisted of an undamped, non-contractile elasticity in series with a damped elasticity that represented the contractile element. Levin and Wyman stated that the damped component behaved in a viscous manner. Fenn and Marsh recognised that the properties of the damped or contractile element could be revealed by maintaining a constant force during shortening; in that case, the length of the undamped elastic component is constant and the shortening must reflect the properties of the contractile element. The viscous elastic model predicted that contractile element’s shortening velocity would decrease linearly with increasing load. Using afterloaded contractions to keep the force constant during shortening, Fenn and Marsh determined the force–velocity properties of cat and frog muscles. The force–velocity relationships were not linear but rather had the now familiar curved shape, indicating that muscles could not have a contractile element that behaved in a viscous-elastic fashion. In a superb and forthright summary, Fenn and Marsh (1935) wrote,
It was shown by Fenn [1924] in Hill's laboratory that in order to shorten and do work a muscle must liberate an extra amount of energy more or less in proportion to the work done in shortening. This fact was held to cast doubt upon the theory that a muscle is simply an elastic body. In spite of this objection subsequent authors have continued to explain phenomena observed in shortening muscles on the basis of simple mechanical models. The present experiments show again that these simple models are inadequate for the purpose, and it seems most likely that they are inadequate for the same reason that muscle is not an elastic body.
In 1937, Hill published a methods paper that marked the culmination of 25 years of development of myothermic apparatus that started with his first Bürker-derived system (Hill 1913b). Hill (1937) described a new fast-responding, low heat capacity thermopile linked to a fast but sensitive galvanometer. With this system, the heat recording step response was complete in ~ 200 ms, a 25-fold increase in speed since Fenn’s studies (line “D”, Fig. 5). The thermopile was equipped with a “protecting region”, an extra section of thermopile under the end of the muscle at which the lever was attached. This was not part of the temperature recording circuit but served to ensure that heat loss from the muscle into the thermopile was the same along the part of the muscle over the recording region at the start of the contraction and along the length of muscle that moved onto the thermopile during shortening. This simple measure solved the long-standing problem that increases in heat during shortening may have been due to warmer sections of muscle moving onto the thermopile (Hill 1913b). Hill had also further developed the deconvolution analysis to correct for both lag in heat records due to the time taken for heat to move from the muscle to the thermocouples and for heat lost from the muscle through the thermocouple wires. Finally, a much-improved heat calibration method had been developed.
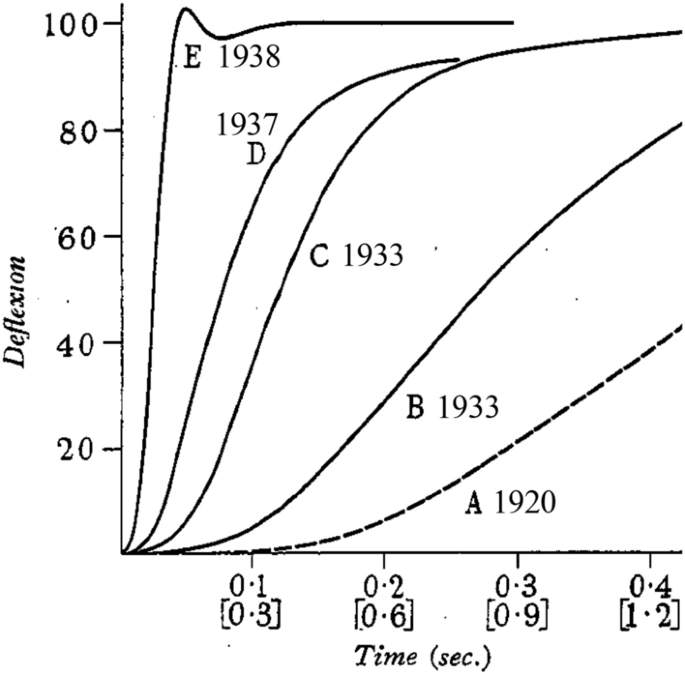
In a report to The Physiological Society, UK, in May 1938, Hill (1938a) gave a preliminary description of experiments, performed using the new apparatus, that unequivocally confirmed Fenn’s conclusions, unveiled Hill’s mathematical description of the force–velocity relation and illustrated that muscle might usefully be considered as consisting of a contractile element, with the observed force–velocity properties, connected in series with an elastic element. This work was published in full in October in a paper entitled The heat of shortening and the dynamic constants of muscle (Hill 1938b). Hill also presented the results to the Physical Society in November and he recommended (Hill 1965b) the published version of that presentation (Hill 1939) as “more readable than the original paper”. An example of Hill’s continual pursuit of better methods was that between the work described in May and the full paper in October, an even faster galvanometer system had been developed, giving the system a step response time of just 50 ms (Fig. 5) (for a description, see Hill 1939).
With this apparatus, Hill concentrated on recording muscle heat production in tetanic contractions in which the muscles were initially held isometric and then at a specified time were allowed to shorten a set distance against a constant load. In planning the experiments, Hill adopted the constant force protocol described by Fenn and Marsh (1935) so that the observed force–velocity properties could be attributed solely to the contractile element. The results of the experiments were clear: the rate of heat production increased abruptly above the isometric rate when shortening started and decreased when shortening ended (Fig. 6A). The amount of heat produced was largely determined by how much the muscle shortened and was not greatly affected by the load nor by the time within the contraction that shortening started (Fig. 6A). In Fig. 6B a summary is shown of the dependence of rate of energy output on the velocity of shortening for one muscle. Work output is 0 in an isometric contraction and when shortening at maximal velocity (Vmax) and rises to a maximum at about 25% Vmax. The rate of heat output increased with shortening velocity, as did the rate of energy output, as both heat and work. This was a convincing demonstration that extra energy was mobilised as the muscle shortened and performed work and that the extent of the mobilisation depended primarily on how much shortening occurred. Although this conclusion was the same as Fenn’s, the heat records with the greatly improved time resolution provided unequivocal evidence. And thus, after almost 100 years, the elastic theories were finally extinguished.
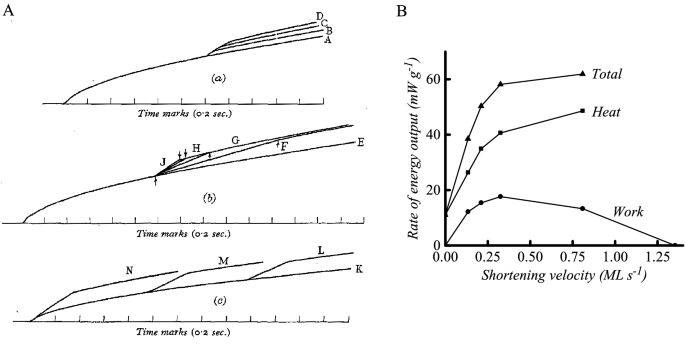
Much of Hill’s discussion about the energy output during shortening focussed on what he called “shortening heat”, which was the heat in excess of the underlying isometric heat. This idea was problematic, with considerable discussion over what constituted the appropriate baseline (Hill 1964; Mommaerts 1970; Rall 1982). Hill (1939) did suggest that it might be more accurate to think of the increased rate of heat output as due to a change in the rate of all the heat producing processes involved in generating force rather than just the addition of an extra component, a view that is closer to our current understanding than the shortening heat concept. However, the scope for mechanistic thinking in 1938 was limited, given that the scheme of chemical reactions occurring in muscle was not known, the distinction between the events of activation and those of force generation had not been elucidated and the uncovering of the structural basis of force generation was still 20 years in the future.
The 1938 paper wasn’t Hill’s last word on the topic. After time spent on war-related activities (for descriptions, see Barclay and Curtin 2022; Katz 1978; Rall 2017), Hill returned to science in the late 1940s. Following further developments of his heat-measuring apparatus (Hill 1949a), Hill showed that more energy, as both heat and work, was produced in twitches with shortening (Hill 1949b, 1953), correcting an earlier anomalous result (Hartree and Hill 1928a). Further, when Hill and others repeated his 1938 experiments with improved apparatus, the relationship between shortening heat and distance shortened was not as simple as proposed in 1938 but instead showed a degree of load-dependence (Hill 1964). This result does not diminish the idea that performance of work and production of heat are related but did make the relationship between the mechanical performance and energy output less straightforward than suggested in 1938.
There was limited scope for insights into the basis of the increase in heat output that accompanied the transition from isometric contraction to work generation, particularly in 1938 but also in 1964 when Hill completed his revisionary experiments on heat output from muscles performing work. However, in the 20 years after the 1964 paper, several key aspects of muscle energetics were elucidated. These are briefly summarised in the following paragraphs; Rall (2018) gives a comprehensive account of these and many other aspects of muscle physiology.
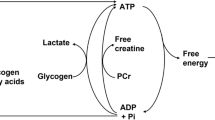
Although it had been recognised since at least the 1840s that chemical reactions must ultimately account for muscle heat production, knowledge of the nature of the reactions that occurred as a muscle contracted remained fragmentary prior to the 1960s. For 30 years after the discovery of ATP in muscle (Fiske and Subbarow 1929; Lohmann 1929), there was only indirect evidence that it played a primary role in contraction. The only consistent chemical change seen to occur in muscle within the time course of a short contraction was a decrease in PCr concentration (for reviews, see Mommaerts 1969; Needham 1950). In 1962, Davies and colleagues provided direct evidence that ATP splitting was the primary energy-providing reaction in muscle (Cain and Davies 1962; Cain et al. 1962). They first showed that under normal circumstances in frog muscle, PCr was broken down in proportion to the amount of work a muscle performed. They then inhibited the creatine kinase (CK) reaction (Reaction 2, Table 2) using dinitrofluorobenzene (DNFB), preventing phosphorylation of ADP by PCr; in response to stimulation, the muscles did contract but there was no decrease in PCr concentration. The concentration of Pi increased, as in untreated muscle, but now ATP concentration decreased and the concentrations of ADP and AMP increased (Reactions 1 and 4, Table 2). This established the currently accepted reaction scheme: ATP splitting provides the energy for contraction and ADP formed in that reaction is rapidly rephosphorylated at the expense of PCr in the cytoplasm (i.e. the CK reaction; Reaction 2, Table 2). PCr is subsequently regenerated using ATP in a reversal of the CK reaction (Reaction 6), that occurs in the mitochondrial intermembrane space proximal to the adenine nucleotide translocase and is catalysed by a different version of CK, using ATP produced by oxidative or anaerobic processes (Reactions 7 and 8, Table 2).
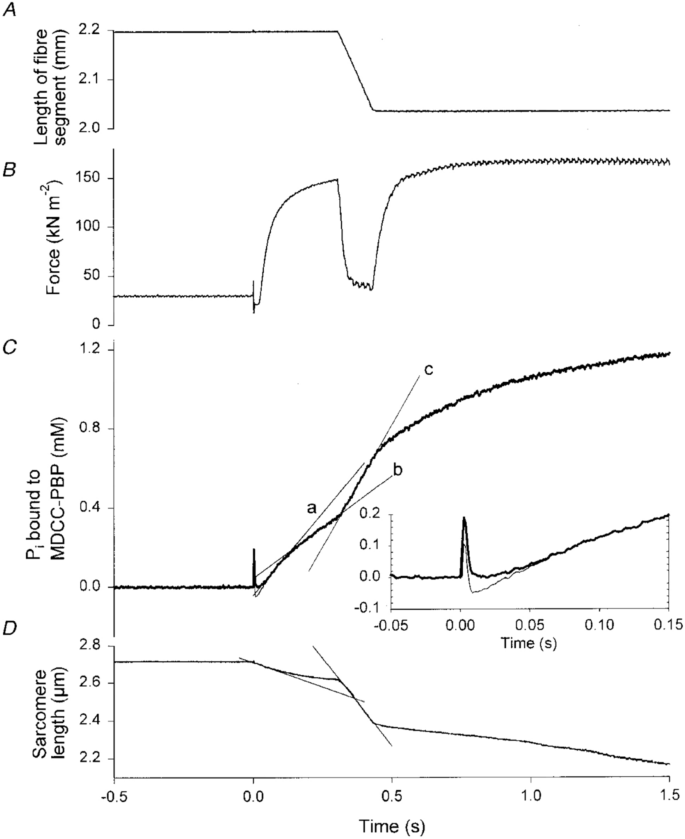
The time course of Pi formation is reminiscent of Hill’s heat records and it is reasonable to take this as confirmation that the change in rate of heat output at the onset of shortening in Hill’s experiments was due to an increase in the rate of ATP splitting by cross-bridges. The Ferenczi group’s experiments provide direct confirmation that a shortening muscle mobilises energy, from ATP splitting, as and when it is required to perform work, as proposed so clearly, but with limited evidence, by Fenn 76 years earlier.
Data from frog sartorius (Hill 1938b) and rabbit psoas (He et al. 1999) already described in this article are consistent with the idea that ATP turnover alters dynamically depending on the mechanical loading encountered during a contraction. But is this a characteristic of all skeletal muscles? The abrupt increase in rate of heat production when shortening starts has been shown to occur in muscles from tortoise (Woledge 1968), fish (Curtin and Woledge 1991) and mice (Barclay et al. 1993, 2010b). Increased rate of Pi release has been observed not only for rabbit muscle but also for fast and slow fibres from human muscle (He et al. 2000). The human fibre study is important because all prior investigations had used muscles or fibres from small animals. However, the human muscle fibres exhibited faster ATP turnover during shortening than during isometric contraction, just as seen for muscles from other species (Fig. 8). In several experiments, the effect of shortening on rate of ATP splitting in permeabilised fibres was measured using an assay in which ATP hydrolysis is linked to the oxidation of NADH, a reaction which can be followed spectrophotometrically (Glyn and Sleep 1985). The temporal resolution of the NADH-based assay is usually not as good as that of the Pi probe (with the exception of Sun et al. 2001) but the overall effect of shortening on ATP use can be calculated. Using that approach, faster ATP hydrolysis during shortening has been demonstrated in rabbit psoas fibres (Potma and Stienen 1996; Sun et al. 2001) and in different fibre types of rat muscle (Reggiani et al. 1997).
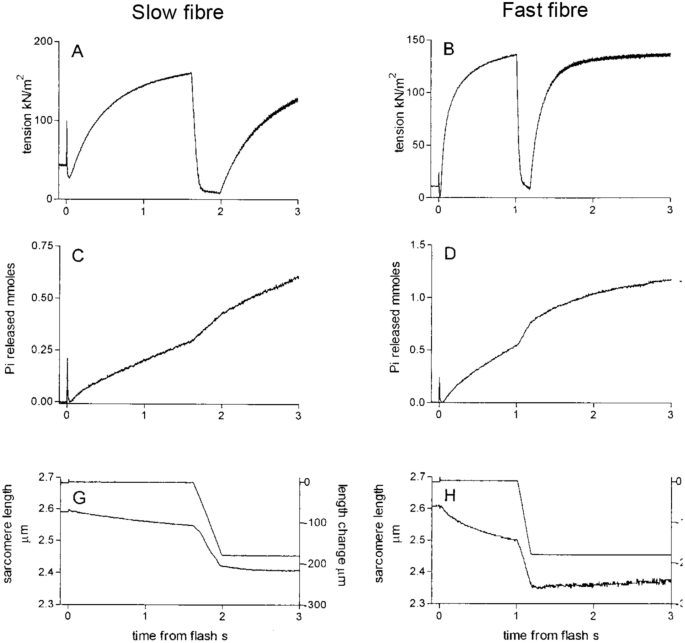
Overall, it does appear that dynamic mobilisation of energy as it is required is a fundamental characteristic of skeletal muscle. It is worth recognising some differences between permeabilised preparations and intact muscle fibres. Permeabilised fibres are provided with simplified energy supply systems and the metabolic factors that limit ATPase activity in permeabilised fibres may differ from those in intact muscle (West et al. 2004). For example, although the Pi probe provides a direct view of ATP splitting, it also prevents the accumulation of Pi (He et al. 1999). In intact muscle, Pi accumulation slows the rate of ATP splitting (West et al. 2004). Conversely, although heat measurements are non-specific (i.e. heat may arise from multiple sources), when used with the protocol guidelines already mentioned, they provide a good index of ATP turnover subject to normal physiological constraints.
An indirect confirmation of the validity of the dynamic energy mobilisation concept is provided by biomechanical modelling studies of the movement of human limbs. In most multi-muscle models of limb movement, each muscle’s behaviour is modelled using an extended version of Hill’s two-component muscle model. That is, the contractile element is characterised by a force–velocity relationship which allows the muscle’s force output at any moment to be determined from the velocity at which the muscle is shortening or lengthening. Account is taken of the time course of activation, prior activity, etc (Winters 1990). Some of these models incorporate muscle energetics. In that case, the contractile element is also supplied with a velocity-dependent heat output (like that shown for frog muscle by the total energy output line in Fig. 6B). Like the force–velocity relation, this is the steady-state relationship determined in the same way as described by Hill (1938b) and scaled appropriately to human muscle (Tsianos et al. 2012; Umberger et al. 2003). The energetics predicted by such models align well with measured human energy output (Tsianos and MacFadden 2016; Tsianos et al. 2012; Umberger et al. 2003). It is particularly notable that even when using steady-state energetic profiles, these models can successfully predict energy use in rapid movements (Umberger et al. 2003). This emphasises that the energetic response of muscles to activity is rapid and dynamic. The success of such models indicates that the basic energetic properties obtained using isolated muscle preparations are applicable to complex real-world muscular activity.
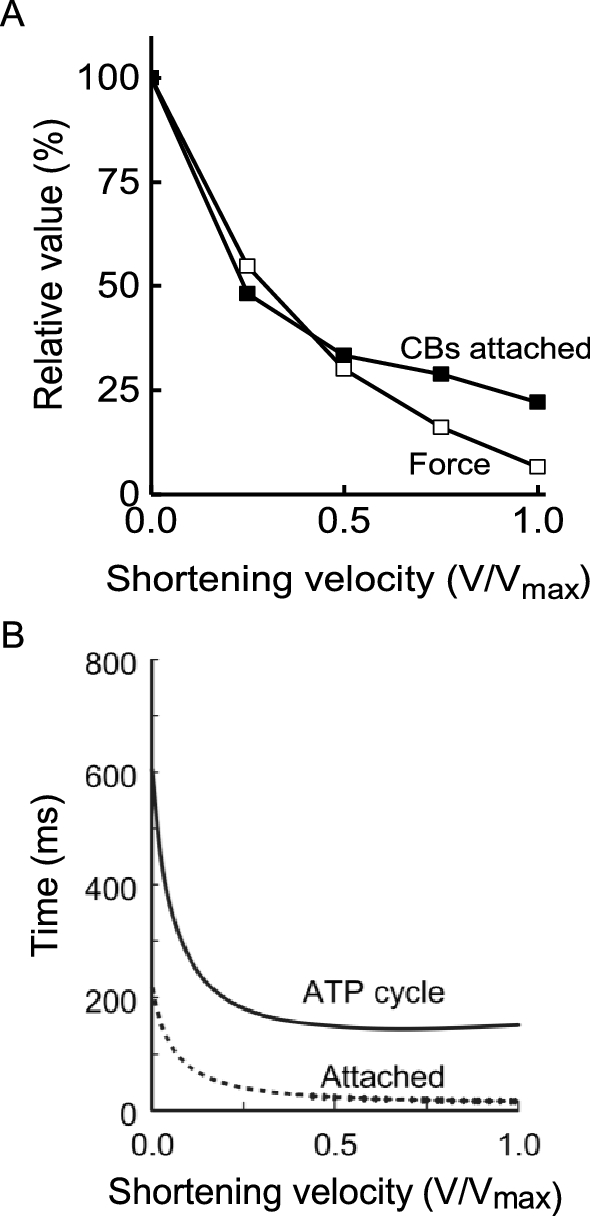
Measurements of the rate of cross-bridge ATP splitting, determined from either Pi probe data or from energy output, can be used to gain insight to the changes in cross-bridge behaviour that produce the shortening velocity dependence of rate of ATP turnover. In a contracting muscle, cross-bridges go through a mechanical cycle involving attaching to a binding site on actin, generating force that promotes filament sliding and detaching. The mechanical cycle is linked to a biochemical cycle with, in its simplest form, a cross-bridge binding ATP prior to attachment and ATP hydrolysis and sequential release of the products Pi and ADP during the attached phase. During isometric contraction of frog muscle, between 30 and 40% of all cross-bridges are attached simultaneously (for explanations of how this can be estimated, see Barclay et al. 2010a; Linari et al. 2007; Offer and Ranatunga 2010). Cross-bridges that are not attached are presumably in the detached part of their cycle, repriming before attachment is again possible. When a fibre is actively shortening, fewer cross-bridges are attached than in an isometric contraction (Fig. 9A) (Barclay et al. 2010a; Caremani et al. 2015; Ford et al. 1985; He et al. 1999). This indicates that during shortening the repriming part of the cycle occupies a larger fraction of the full cycle than in an isometric contraction (Barclay et al. 2010a). However, the increased rate of ATP splitting indicates that the time to complete a full cycle has also decreased (Fig. 9B). Therefore, during shortening cross-bridges cycle faster and spend less of their cycle attached and the overall rate of ATP splitting (the product of cross-bridge cycling rate and cross-bridge concentration) is greater than during isometric contraction. The mechanism by which shortening influences the duration of cross-bridge attachment must involve rate constants for transitions between attached states that are sensitive to the mechanical strain experienced by the cross-bridge. Elucidation of the exact mechanism remains an active area of research (Caremani et al. 2015; Homsher et al. 1997; Linari et al. 2015).
Fenn’s 1923 and 1924 papers are now viewed as a landmark in the development of ideas about muscle energetics. However, it would be fair to say that his work was not decisive; the limitations of the experimental methods made the results a less than reliable basis for the final interpretation. Fenn’s clear expression of his conclusions probably transcended the results of his experiments. But he found independent support for his ideas from his force–velocity experiments in 1935 and, perhaps most importantly, his work stimulated further research. Although Hill’s prolonged adherence to the elastic theory was a major element in the delay in accepting Fenn’s ideas, he deserves credit for recognising what was required to settle the matter and persisting for 15 years with technical developments to the point where he could show that his earlier ideas were wrong and that Fenn was right.
To understand Fenn’s results, and those of Hill, required the surge in knowledge that occurred after Hill’s final work on the matter in 1964. That era provided knowledge of how force and work was generated in muscle and how muscle biochemistry was related to both heat measurements and the force-generating cross-bridge cycle. That knowledge provided a basis for understanding the “Fenn effect” as a reflection of accelerated ATP splitting by cross-bridges when filament sliding commenced, promoted by strain-dependent rate constants for steps within the attached phase of the cross-bridge cycle (Fig. 10). Assembling that mechanism can be considered as the final stamp of approval for Fenn’s ideas, a process 76 years in the making. Fenn’s bold pronouncement of the implications of his work for understanding muscle was ultimately proved right and marked out a key concept in muscle energetics:
The rate at which energy is liberated from ATP hydrolysis by cross-bridges during the contraction is modulated by the loads encountered during the contraction.
Open Access funding enabled and organized by CAUL and its Member Institutions.
